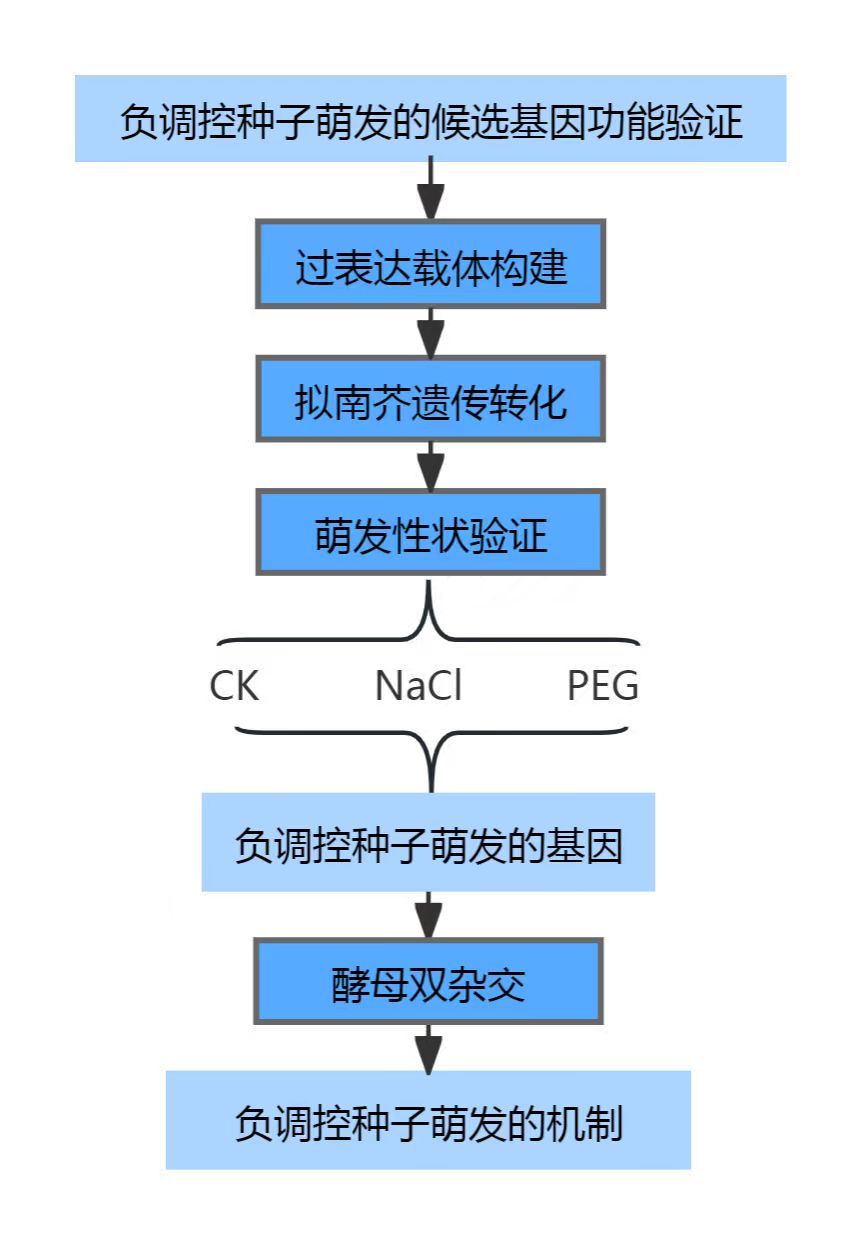
1.1大豆种子萌发研究基础
前期在研究大豆种子萌发试验过程中发现,不同大豆品种发芽速率显著不同,1041个大豆品种中有7个品种24 h内全部发芽,有52个品种24 h内不发芽。萌发速率影响苗的强壮和后期植株的生长,有必要探明影响大豆发芽速率的机理机制,以便于控制大豆种子萌发进程,提高大豆产量。
试验以绥农28为材料,将大豆种子萌发细分为三个不同阶段,发现在0-18 h 种子快速吸涨,吸水平均速率为13.1 mg?h-1?粒-1,这是大豆种子吸涨阶段。在18-36 h大豆种子重量只有较小的变化,吸水速率平均仅为1.0 mg?h-1?粒-1,这个阶段定义为大豆胚根萌动阶段,当大豆胚根突破种皮后,定义为胚根伸长阶段。
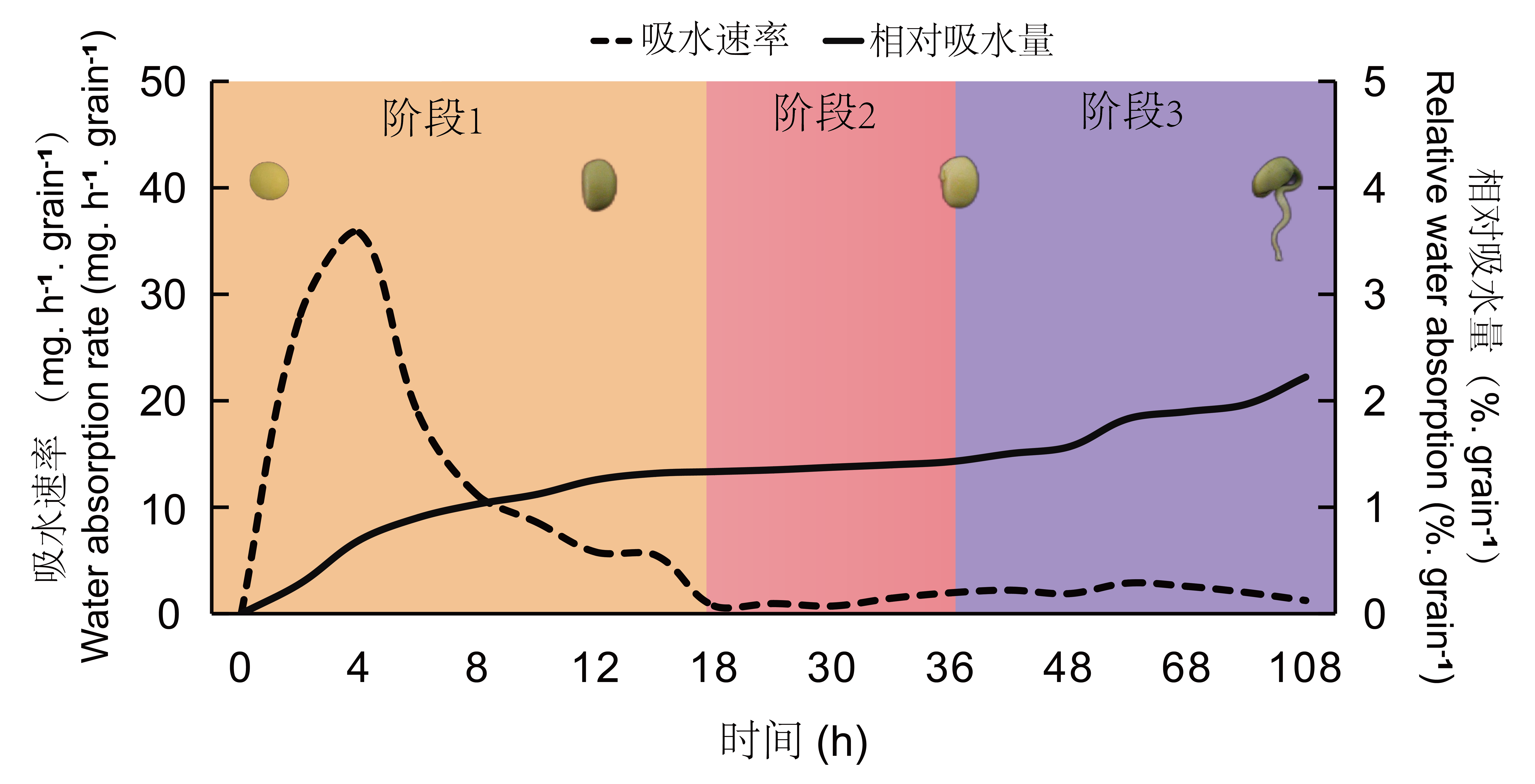
图1 绥农28种吸水速率
1.2 大豆种子萌发时mRNA转录组分析
以大豆品种绥农28为材料,分别在浸种12h(种子吸涨)、36h(胚根萌发)和108h(胚根伸长)研究大豆胚芽中mRNA表达谱,种子萌发时差异表达的基因有20845个,对差异基因进行基因趋势分析,发现同一mRNA在同一个组织的不同时间点时表达量具有时序性。
为了探讨不同时间点差异基因的功能,根据表达趋势对 20854 个差异基因进行分类,得到 8 个趋势(P1-P8)。P1 和 P6 包含超过 43.77%的差异基因,在 12 h VS 36 h 和 36 h VS108 h 两个阶段趋势一致,表明这些基因即参与种子吸胀萌发过程也控制胚根伸长过程。P2 和 P5 包含 34.91%的差异基因,这些 DEM 在 12 h VS 36 h 中具有差异,但在 36 h VS 108h 变化较小,意味着它们可能与种子萌发突破种皮关系更大。P3 和 P4 包含超过 16%的基因,在 12 h VS 36 h 阶段变化较小,在 36 h VS 108 h 具有差异,这意味着这些 DEM 可能与种子萌发突破种皮关系较小,但和胚根伸长关系更大。P7 和 P8 所包含的基因数量最少,且在 12 h VS 36 h 发生变化后,在 36 h VS 108 h 分别发生回调,意味着他们可能仅与胚根突破种皮这一过程有关(图2 )。
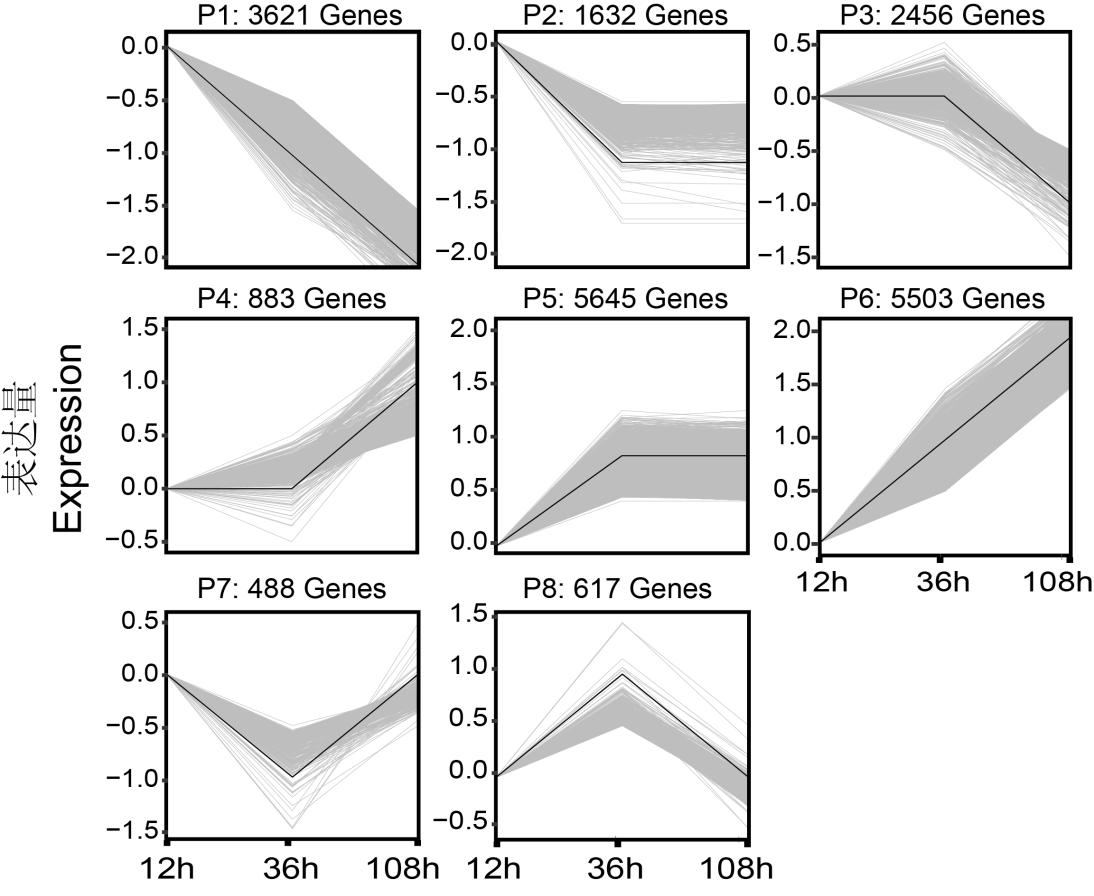
图2 差异基因趋势分析。
1.3 差异 mRNA(DEMs)功能分析
为了进一步 8 种 DEM 趋势在功能富集上的异同,我们对 8 个趋势中的 DEMs 分别进行 KEGG 富集分析,其中 P1 和 P4 主要参与“植物激素信号转导 (ko04075)”、“植物-病原体相互作用(ko04626)”和“剪接体(ko03040)”。P2 主 要富集在“剪接体(ko03040)”、“RNA 转运(ko03013)”和“内质网中的蛋白质加 工(ko04141)”。P3 主要富集在“氨基酸的生物合成(ko01230)”、“植物-病原体相 互作用(ko04626)”和“植物激素信号转导(ko04075)”。P5、 P6 和 P7 主要参与“植 物激素信号转导(ko04075)”、“植物-病原体相互作用(ko04626)”和“MAPK 信号 通路(ko04016)”。P8 主要富集在“植物激素信号转导(ko04075)”、“植物-病原体相互作用(ko04626)”和“核糖体(ko03010)”。(图 3-4 A)。 通过对 COG_class_annotation 的分析,我们发现 P1 和 P4 主要具有翻译后修饰、信 号转导等功能,P2 主要具有翻译及翻译后修饰功能,P3 主要具有翻译后修饰和碳水化合 物代谢功能,P5 主要具有一般功能,P7 主要具有信号转导和碳水化合物代谢功能,P8 主 要具有碳水化合物代谢和一般功能(图 3)。
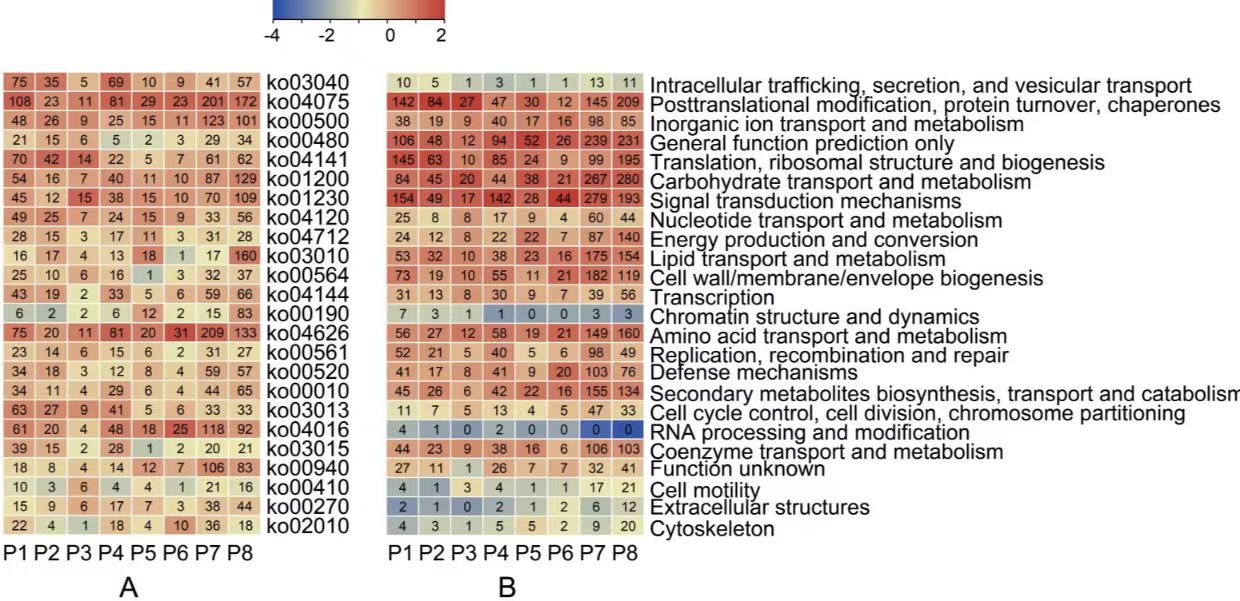
图 3 DEMs 功能分析
A:KEGG 通路富集;B:8 个趋势 COG 类注释
1.4 mRNA的确定
为此我们挑选属于P1趋势的17个差异基因,包括COL9 (zinc finger protein CONSTANS-LIKE 13-like)、LOC100809693 (translation factor GUF1 homolog, chloroplastic )、LOC100816920 (aldehyde dehydrogenase 22A1 )、LOC100787296 (DNA polymerase V)、LOC100795995 (trihelix transcription factor GTL1)、LOC100811164 (ABC transporter G family member 50、LOC100779103(protein MLN51 homolog)、LOC100817761 (pre-mRNA-splicing factor ATP-dependent RNA helicase DEAH7)、LOC100812693 (APO protein 3, mitochondrial)、LOC100801868 (trihelix transcription factor GTL2)、LOC100806616 (RIK protein )、LOC102664661 (ankyrin repeat protein SKIP35)、LOC100804754 (AMSH-like ubiquitin thioesterase 1)、LOC100796687 (SNF1-related protein kinase catalytic subunit alpha KIN10)、LOC100809289 (vacuolar protein sorting-associated protein 35A)、LOC100810186 (beta-(1,2)-xylosyltransferase isoform X1)、LOC100812846 (F-box protein CPR1)。这17个基因对大豆种子萌发的抑制功能,需要通过试验方法去验证。
表1. 基因KEGG途径及差异倍数信息
基因号 | KEGG途径 | 12hVS36h DESeq2_log2FC | 12hVS108h DESeq2_log2FC |
COL9 | ko04712 | -1.907352482 | -3.666186014 |
LOC100809693 | -- | -1.734187648 | -3.104099594 |
LOC100816920 | ko00650 | 0.481874148 | -3.022110566 |
LOC100787296 | -- | -1.425958596 | -2.760727114 |
LOC100795995 | ko00040 | -1.219836634 | -2.394000301 |
LOC100811164 | ko02010 | -0.28847551 | -2.250134526 |
LOC100779103 | -- | -0.806090372 | -2.132992981 |
LOC100817761 | ko03040 | -0.851623157 | -1.91890249 |
LOC100812693 | -- | -1.452827136 | -1.80752683 |
LOC100801868 | ko00040 | -1.423492381 | -1.66242658 |
LOC100806616 | -- | -0.952716323 | -1.544577129 |
LOC102664661 | -- | -0.79004497 | -1.224609886 |
LOC100804754 | ko04144 | -0.896456952 | -1.142508125 |
LOC100796687 | -- | -0.932545334 | -1.036558092 |
LOC100809289 | ko04144 | -0.472943861 | -1.716586043 |
LOC100810186 | ko00513 | -1.489926639 | -3.173481431 |
LOC100812846 | -- | -0.793459961 | -1.079635408 |