2024
肠瘟净---一种新型水貂二联疫苗
盲选
创新训练项目
农学
动物医学类
学生来源于教师科研项目选题
一年期
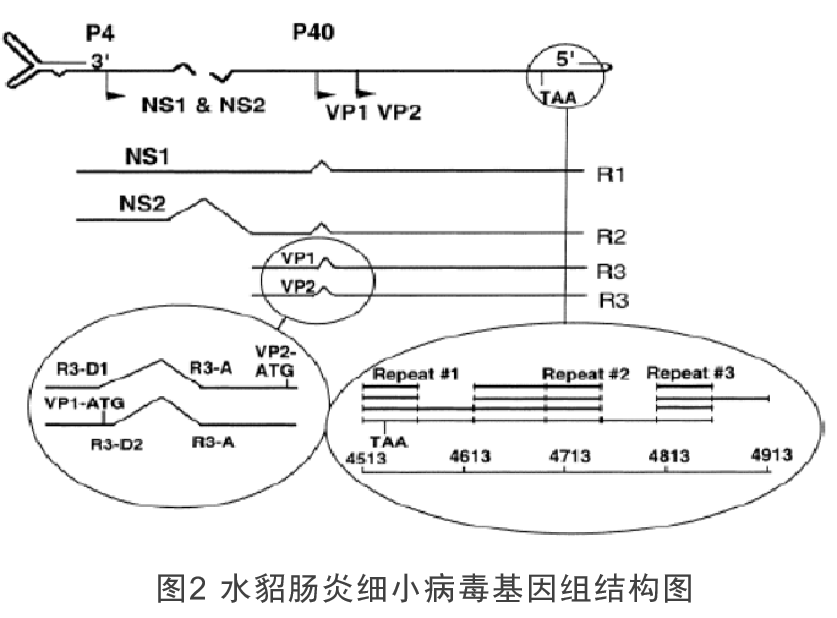
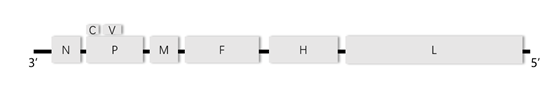
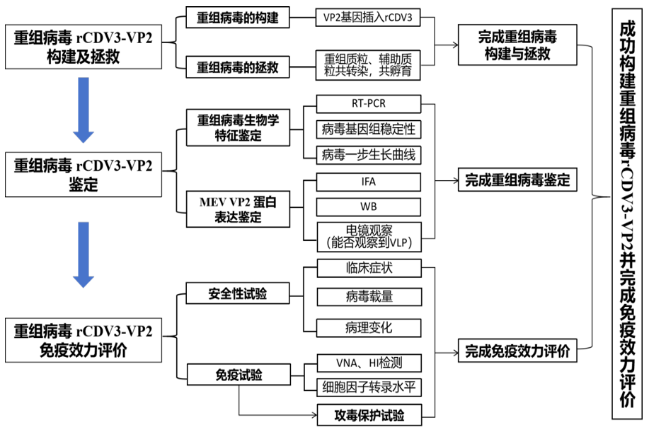
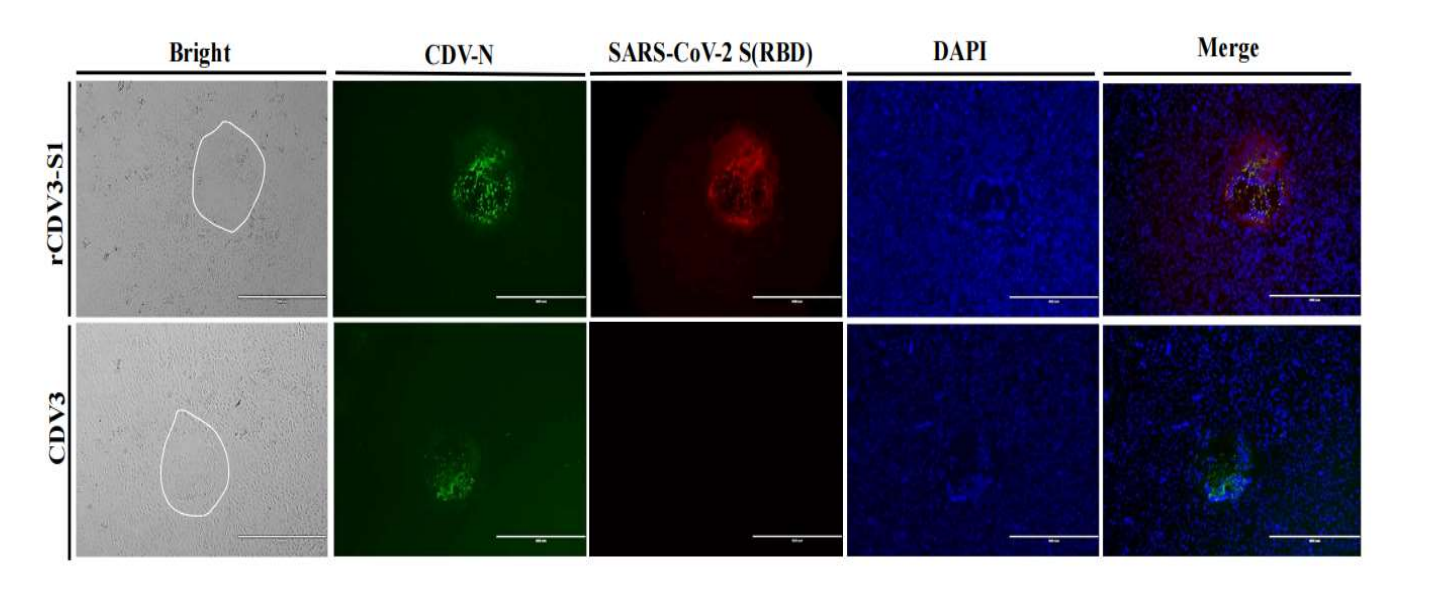
为应对水貂养殖业的需求,本项目拟利用反向遗传技术以水貂犬瘟热病毒弱毒CDV3株的拯救为基础,在CDV基因组P-M中克隆入MEV VP2基因,经过病毒拯救,获得同时表达CDV蛋白和MEV VP2蛋白的重组犬瘟热病毒rCDV3-VP2株为犬瘟热和水貂细小病毒性肠炎二联苗的研发提供候选疫苗株。
1、项目负责人已在犬瘟热课题组一进行了一年的学习,熟练掌握相关实验的操作原理及过程。
2、负责人参与犬瘟热课题研究一项。
1.国家自然科学基金面上项目,人SLAM受体介导犬瘟热病毒跨种间感染的分子机制,2020年01月至2023年12月;
2.国家自然科学基金青年基金,犬瘟热病毒变异株对水貂致病性增强的分子机制,2016年01月至2018年12月;
3.吉林省重点科技成果转化项目,水貂犬瘟热活疫苗产业化开发,2014年01月至2016年12月;
4.国家重点研发计划子课题,水貂、狐、貉犬瘟热-细小病毒性肠炎二联活载体疫苗创制,2023年12月至2027年12月;
5.黑龙江省重点研发计划项目,畜禽重要传染病快速诊断关键技术研发与推广,2021年9月至2025年08月;
1、主要为项目的研究背景和研究设计提供方向性指导,在难点和重点部分进行把关,帮助项目团队解决项目难题。
2、主要为项目团队的实践创造条件,根据项目团队的实际需求,可以为项目团队提供实地考察的机会。
校级